
Fragmentary specimens are something paleontologists deal with every day: an isolated tooth, a few limb bones, or a jaw with only one or two teeth. Thanks to the vagaries of the fossilization (and discovery) processes, this is often all that remains of an animal that died millions of years ago. Even though fragmentary specimens are quite useful, we paleontologists generally long for more complete specimens: a pristine jaw, a well-preserved skull, or even an entire skeleton. I wish I had a nickel for every scientific paper that suggested the study “must await the discovery of more complete remains” to reach a definitive conclusion!
Sometimes, however, more complete fossil remains aren’t the panacea they are made out to be. A great example of this is the extinct South American caviomorph rodent Neoreomys australis.
Neoreomys australis was named by Florentino Ameghino more than 130 years ago. It was a largish rodent, weighing some 3-5 kg (6-11 lbs.), similar to a modern woodchuck (Marmota monax). It had large, complex, very high-crowned (hypsodont) teeth and well-developed chewing muscles that together suggest it primarily fed on tough, abrasive vegetation relatively close to the ground. Its limbs seem to have been optimized for quick and efficient movements rather than for digging or climbing, though it was probably much less speedy than some modern species such as the Patagonian mara (Dolichotis patagonicum).
Neoreomys was quite abundant in southern South America during the early Miocene, about 18-16 million years ago; its remains are among the most common fossils at sites like Santa Cruz in southern Argentina and Pampa Castillo in southern Chile. Consequently, much of its anatomy is known in detail; complete skulls and many jaws have been collected in addition to a variety of bones of the skeleton.
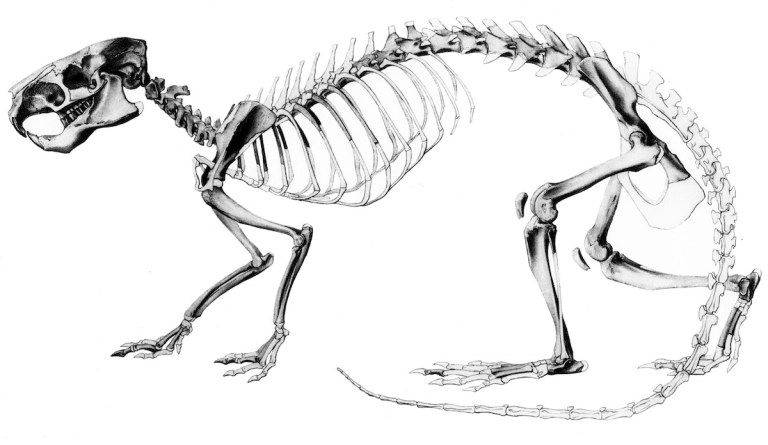
For much of the 20th century, Neoreomys was placed in the family Capromyidae, which encompasses distinctive West Indian rodents known as hutias. Hutias are mostly of medium to large size with varied diets (depending on the species), and some hutias are equally at home in the trees as well as on the ground. Many species of hutias became extinct after humans arrived to Caribbean islands, and fewer than half of the species that were living there about 12,000 years ago are still alive today. Most of those that survived are in danger of extinction. Neoreomys was referred to this family due to the influential work of Wiliam Berryman Scott (1905), who described many well-preserved remains of Neoreomys from Santa Cruz and noted certain aspects of its teeth and skull that resembled capromyids.

Nevertheless, Scott also noted some resemblances between Neoreomys and other caviomorph rodents. In fact, he concluded his description with this statement:
“While [Neoreomys] resembles the [Capromyidae] in dentition and skull structure rather more than any other Recent rodents, it also has many likenesses to Agouti [Cuniculus], Dasyprocta, and Dinomys. It is exceedingly difficult, if not quite impracticable, to determine which of these similarities are due to genetic relationship and which of them to convergent evolution. On the whole, it seems probable that in Neoreomys we have the somewhat modified remnant of a more ancient group, which was the common ancestor of the three families indicated.”
Clearly Scott wasn’t entirely convinced of how to classify Neoreomys.
Ideas about the relationships of Neoreomys began to change in the late 1950s, when Albert Wood and Bryan Patterson suggested that pacas were its closest living relatives. This was based on their detailed study of more ancient rodent skulls from southern Argentina (from the late Oligocene epoch, about 27-25 million years ago). A subsequent study of theirs (published more than 20 years later) that focused on similarly ancient rodent skulls from Bolivia reinforced their initial interpretation. Thus, Neoreomys was transferred to the family Dasyproctidae.
Today, the family Dasyproctidae is recognized as including about a dozen species of rodents knowns as agoutis and acouchis (or acuchis). I like to describe agoutis and acouchis as “souped-up” guinea pigs: medium-sized (up to 4 kg or 9 lbs) tropical rodents with long legs, short tails, and a rump that looks like it is jacked up higher than their head. They are built for speed, at least for a rodent, and spend their time feeding on low vegetation and hunting fruits and seeds that have fallen to the ground. Like Northern Hemisphere squirrels, agoutis and acouchis will collect and bury seeds when they are abundant and return to eat them later.

But what about pacas? If you were paying close attention, you’ll recall that Wood and Patterson thought Neoreomys was related to pacas, not agoutis and acouchis. The issue is that all three types of rodents – pacas, agoutis, and acouchis – were once grouped together in the family Dasyproctidae, which is named for the agouti genus Dasyprocta. The consensus opinion changed in the 1980s with the proliferation of molecular phylogenetic analyses: pacas are now recognized as a separate evolutionary branch and have been shifted to their own family, Cuniculidae.
What they heck are pacas? They include two species of rodents that are much larger than agoutis and acouchis (up to 15 kg or 33 lbs) and less cursorial (speed-adapted). Pacas are also apparently quite good to eat. An immediately recognizable feature of a paca its fur, which is dark with light spots and streaks like that of a young deer. (Check out this nice video of pacas in the wild.) But a paca’s most distinctive feature (unique among mammals) are its cheek bones (zygomatic arches), which are expanded to create hollow resonating chambers for the low, rumbling vocalizations it produces. Pacas were once placed in the genus Agouti (not to be confused with the common name of their smaller relatives discussed above!), but Cuniculus is now considered the proper name for their genus.
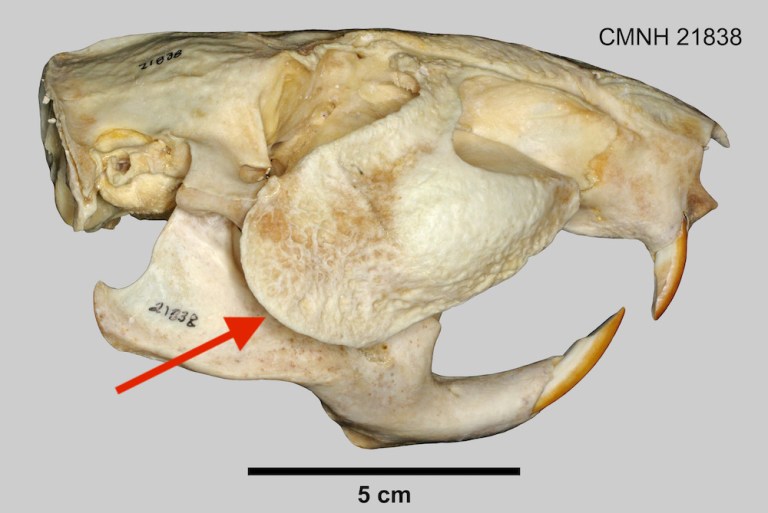
How is Neoreomys classified today?
Most paleontologists place Neoreomys in or near the family Dasyproctidae, but this appears to be due to taxonomic inertia more than anything else; no one has yet done a detailed study of Neoreomys to determine whether it shares unique features with agoutis (and therefore should be kept in the family Dasyproctidae) or pacas (implying it should be shifted to the Cuniculidae) or whether it represents an evolutionary branch separate from both that might even warrant recognition as a new family, Neoreomyidae.
Why are the relationships of one of the best-known early Miocene rodents still virtually unknown? Rodents are a particularly challenging fossil group to work on; there are many species, and convergent evolution is common, especially in their teeth. As a consequence, there are many more rodent species to be studied than there are people studying them. We have plenty of outstanding fossil remains of Neoreomys. What we lack is a Neoreomysologist to study them!
References
- Candela, A. M., L. L. Rasia, and M. E. Pérez. 2012. Paleobiology of Santacrucian caviomorph rodents: a morphofunctional approach; pp. 287-305 in S. F. Vizcaíno, R. F. Kay, and M. S. Bargo (eds.), Early Miocene Paleobiology in Patagonia: High-Latitude Paleocommunities of the Santa Cruz Formation. Cambridge University Press, Cambridge.
- Fabre, P.-H., J. T. Vilstrup, M. Raghavan, C. Der Sarkissian, E. Willerslev, E. J. P. Douzery, and L. Orlando. 2014. Rodents of the Caribbean: origin and diversification of hutias unravelled by next-generation museomics. Biology Letters 10:20140266.
- Lima, S. G. C., R. S. Sousa-Lima, R. S. Tokumaru, S. L. G. Nogueira-Filho, and S. S. C. Nogueira. 2018. Vocal complexity and sociality in spotted paca (Cuniculus paca). PLoS ONE 13:e0190961.
- Patterson, B., and A. E. Wood. 1982. Rodents from the Deseadan Oligocene of Bolivia and the relationships of the Caviomorpha. Bulletin of the Museum of Comparative Zoology 149:371-543.
- Scott, W. B. 1905. Mammalia of the Santa Cruz Beds. Volume V, Paleontology. Part III, Glires; pp. 384-490 in W. B. Scott (ed.), Reports of the Princeton University Expeditions to Patagonia, 1896-1899. Princeton University, E. Schweizerbart’sche Verlagshandlung (E. Nägele), Stuttgart.
- Upham, N. S., and B. D. Patterson. 2015. Evolution of the caviomorph rodents: a complete phylogeny and timetree of living genera; pp. 63-120 in A. I. Vassallo, and D. Antenucci (eds.), Biology of Caviomorph Rodents: Diversity and Evolution. SAREM Series A, Buenos Aires.
- Wood, A. E., and B. Patterson. 1959. The rodents of the Deseadan Oligocene of Patagonia and the beginnings of the South American rodent radiation. Bulletin of the Museum of Comparative Zoology 120:282-428.

2 thoughts on “Rodent of Uncertain Systematics (R.O.U.S.)”