
I teach head and neck anatomy to medical students. This part of the body includes a lot of really interesting anatomy, but one aspect that all students find fascinating are the ear ossicles: the three tiny bones that sit in our middle ear and transmit sound from our eardrum to the fluid of our inner ear.
Why are students so impressed by these tiniest of bones? It is probably their combination of small size and complex structure. In humans, one of the bones is shaped like a tiny stirrup (the stapes) yet measures only a few millimeters. It is quite fragile for a bone, with the result that few students are able to extract it in lab without damaging it. The other ossicles – the malleus (hammer) and incus (anvil) – are beefier and less frequently damaged by students, but this does not keep them from getting lost amongst the pieces of temporal bone that must be removed to view them.

The fossilization process is no more forgiving than an anatomy lab. Bones are frequently lost or destroyed, especially the ear ossicles. For that reason, there isn’t a lot written about the ear ossicles of most Cenozoic fossil mammals, at least compared to what is written about less fragile parts like teeth, other portions of the skull, and limb bones. But that doesn’t mean such bones are completely unknown.
I was reading a recent article by Gerry De Iuliis and colleagues on a couple of Pleistocene megalonychid sloths from Brazil, and I was impressed by the beautiful figure they included comparing the size and structure of ear ossicles (malleus and incus) of several species of extinct sloths. After a little digging, I discovered that the study of sloth ear ossicles has a rather long history. The ear bones of modern sloths were first described by Alban Doran in 1878, and only a couple decades later, Woodward was the first to describe the bones of an extinct sloth (Mylodon) in 1900. There haven’t been a lot of studies of sloth ear bones since then, but there have been a few. One standout is a very detailed comparative study begun by Bryan Patterson and Walter Segall in the late 1940s that was ultimately published posthumously by Bill Turnbull and Tim Gaudin in 1992.

Is there any reason to study sloth ear ossicles other than just classic, descriptive anatomy? One of the primary motivations of most researchers studying these tiny structures is to use them to uncover evolutionary relationships. The ear region and other complex parts of the base and back of the skull tend to be quite useful for such studies, but they are particularly useful in this case; sloths and other xenarthrans lack complex teeth, which are a fundamental source of information about evolutionary relationships in most other groups of mammals. As a consequence, sloth teeth have provided comparatively little information about how sloths are related to one another. The ear region and other parts of the sloth skull can help compensate for that.
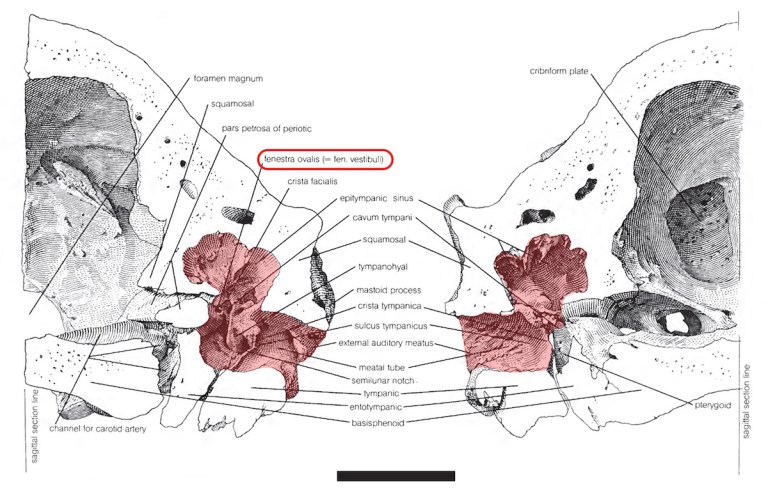
Another reason to study ear bones is to learn something about the hearing capacities of extinct animals. Small mammals are often the object of such studies, but these types of analyses have also been carried out for extinct sloths. In 2008, Ernesto Blanco and Andrés Rinderknecht studied the size of the tympanic membrane (eardrum) and ear ossicles in two large Pleistocene sloths (Glossotherium and Lestodon) from Uruguay. Based on the relative sizes of these structures, they concluded that these sloths may have used low-frequency sound for long-distance communication like elephants do today. Considering that Lestodon was almost as large as an elephant (4 tons or 4,000 kg), this comparison does not seem like too much of a stretch.
Earlier this year, Blanco and Washington Jones published a similar study of eardrum size in a handful of early Miocene (18 million-year-old) sloths from southern Argentina. They found a fair bit of variation among them and divided them into three groups based whether their inferred optimal hearing frequency was lower than, equal to, or greater than that predicted based simply on their body size (mass). They inferred that sloths with low-frequency hearing (Hapalops and Analcimorphus) were nocturnal and/or fossorial, lived in low-density groups, had small external ears (pinnae), and communicated over long distances. By contrast, they concluded that sloths with high-frequency hearing (Pelecyodon and Schismotherium) had better vision, were more arboreal, had larger pinnae, and lived in groups. They inferred that the one sloth with “typical” hearing, Analcitherium, had habits similar to high-frequency species. Blanco and Jones admit that their paleobiological inferences are speculative (one might say highly speculative), and since their study was based on published illustrations rather than the specimens themselves, their data should be taken with a grain of salt. Nevertheless, this type of study at least provides hypotheses that can be tested in the future using other, more precise methods.
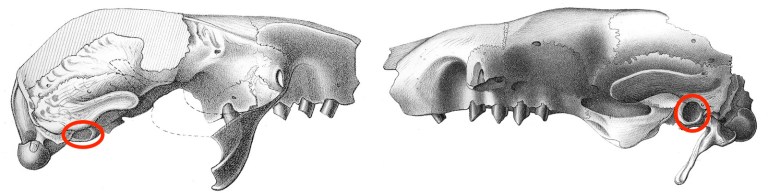
Now that high resolution CT scanners are common, studying the structure of the inner ear – which is completely housed within a blocky bone at the base of the skull known as the petrosal – is much easier and less damaging to specimens than it was previously. The inner ear is where hearing actually takes place and is also responsible for sensing position and motion. Therefore, many investigations of the inner ear aim to better understand animal biology. Several years ago, Guillaume Billet and colleagues published an interesting study showing that the vestibular apparatus of modern three-toed sloths (Bradypus variegatus) – the part of the inner ear that senses position and motion – is more variable than that of other mammals. They reasoned that this was probably due to this sloth’s extremely relaxed lifestyle. Mammals that move quickly need precise and accurate information about their movements, lest they fall off a branch or make a false step while evading a predator. Since sloths are never in a rush, they apparently can do just fine with lower-quality positional data. It would be quite interesting to see if the same was true of extinct sloths given their considerable diversity in shapes and sizes. Unfortunately, measuring such variation requires a large sample size of a single species, which is often difficult to come by in the fossil record.
One thing all these studies have in common – other than ear of sloth, of course – is that they illustrate how new specimens, new approaches, and new technologies can all lead to new insights about living and extinct species. Paleomammalogy isn’t simply about new species and their teeth.
References cited:
- Billet, G., L. Hautier, R. J. Asher, C. Schwarz, N. Crumpton, T. Martin, and I. Ruf. 2012. High morphological variation of vestibular system accompanies slow and infrequent locomotion in three-toed sloths. Proceedings of the Royal Society B: Biological Sciences 279:3932-3939.
- Blanco, R. E., and A. Rinderknecht. 2008. Estimation of hearing capabilities of Pleistocene ground sloths (Mammalia, Xenarthra) from middle-ear anatomy. Journal of Vertebrate Paleontology 28:274-276.
- De Iuliis, G., C. Cartelle, and F. Pujos. 2016. New Pleistocene remains of megalonychid ground sloths (Xenarthra: Pilosa) from the intertropical Brazilian region. Journal of Paleontology 90:578-587.
- Doran, A. H. G. 1878. Morphology of the mammalian ossicula auditus. Transactions of the Linnean Society of London. 2nd Series: Zoology 1:371-497.
- Blanco, R. E., and W. W. Jones. 2016. Estimation of hearing capabilities of Early Miocene sloths (Mammalia, Xenarthra, Folivora) and palaeobiological implications. Historical Biology 28:390-397.
- Patterson, B., W. Segall, W. D. Turnbull, and T. J. Gaudin. 1992. The ear region in xenarthrans (= Edentata: Mammalia). Part II. Pilosa (sloths, anteaters), palaeanodonts, and a miscellany. Fieldiana: Geology (New Series):1-79.
- Scott, W. B. 1904. Mammalia of the Santa Cruz Beds. Volume V, Paleontology. Part I, Edentata. 3. Gravigrada; pp. 227-364 in W. B. Scott (ed.), Reports of the Princeton University Expeditions to Patagonia, 1896-1899. Princeton University, E. Schweizerbart’sche Verlagshandlung (E. Nägele), Stuttgart.
- Woodward, A. S. 1900. On some remains of Grypotherium (Neomylodori) listai and associated mammals from a cavern near Consuelo Cove, Last Hope Inlet, Patagonia. Proceedings of the Zoological Society of London 69:64-78.

Wonderful discovery
LikeLike