
Armadillos are the only living mammals that have a protective armor covering made of skin (dermal) bones known as osteoderms. The only extinct mammals with such a boney covering are close relatives of armadillos such as glyptodonts, which were giant, tortoise-like beasts, and pampatheres, which looked something like a cross between a glyptodont and an armadillo. The most simple explanation for this pattern is that osteoderms (protective skin bones) were present in the common ancestor of all these armored mammals and inherited by all of its descendants, eventually evolving into myriad forms that can be found throughout most of the South American fossil record.
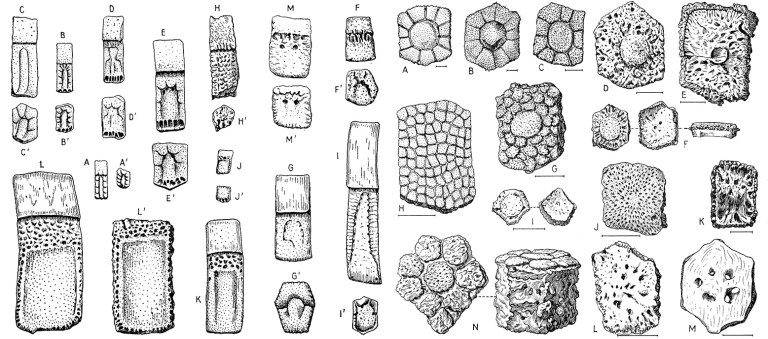
Although the boney shell of armadillos and their extinct relatives (technically known as cingulates) is unique among mammals, cingulates are not the only mammals that had bones embedded in their skin. Extinct sloths of the family Mylodontidae also had such bones, a fact that has been known for more than 150 years.
The presence of osteoderms in an extinct sloth was first noted by Hermann Burmeister, who was director of the natural history museum in Buenos Aires (now the Museo Argentino de Ciencias Naturales ‘Bernardino Rivadavia’). He published a paper in 1865 in which he described a variety of osteoderms associated with a specimen of Glossotherium robustum.* Several decades later, Moreno and Smith Woodward (1899) described a piece of mummified skin from a cave in southern Chile that was riddled with similar structures. The skin was later identified as belonging to the recently-extinct sloth Mylodon darwinii. Since then, osteoderms have been identified in several extinct sloth species from the Pleistocene of Brazil (Mylodonopsis ibseni, Valgipes sp., Ocnotherium giganteum, Glossotherium sp.; Beltrame et al. 2014) as well as two species from the Pliocene and Pleistocene of North America: ‘Glossotherium’ chapadmalense from Florida (Robertson 1976) and Paramylodon harlani from the La Brea Tar Pits (Sinclair 1910), respectively. All of these species belong to the extinct family Mylodontidae, a group of giant sloths that inhabited North and South America until the Pleistocene megafaunal extinction about 12,000 years ago.

Sloth osteoderms do not closely resemble those of armadillos; they lack the consistent pattern of grooves, pits, and other surface structures that are typical of armadillo and glyptodont osteoderms and are rather irregular in shape. Some sloth osteoderms are diamonds or trapezoids, but many resemble small, bony pebbles. Sloth osteoderms are of moderate size, about 5-25 mm (1/4” to 1”) in greatest dimension. They were closely positioned in life, touching one another, but did not form a solid, inflexible shield like armadillo and glyptodont osteoderms.

Thus far, only mylodontid sloths (or mylodonts, for short) are known to have had osteoderms. While it is possible that sloths of other families also had them, this seems unlikely considering that such groups are represented by many well-preserved remains that apparently lack osteoderms. If a Pleistocene sloth like Megalonyx or Megatherium had osteoderms, one would think they would have been discovered by now. But that is part of the fun of paleontology; it only takes one discovery to completely change the story and our understanding of evolutionary history!
Even among mylodonts, it is not known with certainly how many species had osteoderms. The family originated by the late Oligocene epoch, more than 25 million years ago, and the only species identified thus far with osteoderms are relatively young in geological terms (most are thousands rather than millions of years old). One possible explanation for this pattern could simply be the fossil record. Even if most or all mylodonts had osteoderms, the odds of finding small bones that can easily be lost during the fossilization process are much greater for a species that is a mere 12,000 years old than for one that is 12 million years old. This is because there are many more remains of geologically young sloths and they tend to be more complete than those of more ancient species. On the other hand, it could also be the case that osteoderms evolved relatively recently within the family, within the past few million years, perhaps due to some physiological or ecological factor that caused them to be favored by natural selection. It would be easy to fill a page with possible hypotheses for why this might occur, but most of these would be difficult or impossible to test.

Setting aside the question of whether all mylodonts had osteoderms, an important question that probably CAN be answered is how many times osteoderms evolved among mammals. The fact that these structures have only been found in two closely-related groups of mammals – cingulates and mylodonts – has led to the proposition that osteoderms were present in the common ancestor of these two groups and were simply lost in anteaters and non-mylodontid sloths. (Xenarthra is the name of the group that includes sloths, armadillos, and anteaters.) This could certainly be true, but this isn’t the simplest explanation (technically known as the most parsimonious explanation), at least based on our current understanding of xenarthran evolutionary relationships. That’s because it requires three independent losses of osteoderms: in anteaters, three-toed sloths (family Bradypodidae), and the lineage that gave rise to two-toed sloths (family Megalonychidae), the giant megatheres, and many other extinct sloths. In other words, that scenario requires at least four evolutionary steps (one appearance and three losses of osteoderms). A much simpler explanation is that osteoderms evolved twice independently: once in cingulates (armadillos, glyptodonts, and kin) and again in mylodontid sloths.
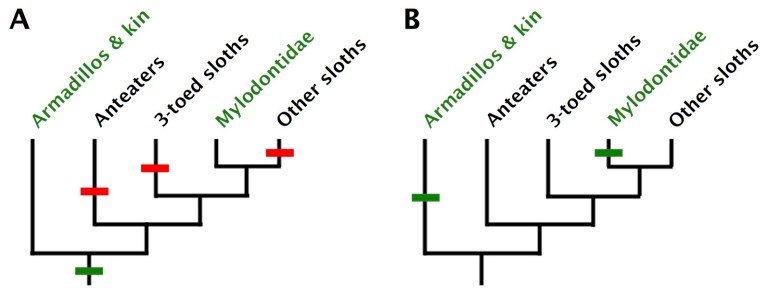
That being said, it does seem coincidental that the only two groups of mammals to ever evolve osteoderms are fairly closely related to one another. Perhaps the common xenarthran ancestor had some propensity for developing these structures that increased the chances of their evolving more than once among its descendants. Or maybe the common xenarthran ancestor really did have osteoderms, and osteoderms just happened to be lost several times independently. Although simplicity (parsimony) is the criterion we use to judge the likelihood of an evolutionary scenario, that does not mean it is right – we just expect (and hope) that it is!
*Burmeister referred to this specimen as Mylodon gracilis, a species now recognized as a junior synonym of Glossotherium robustum (McAfee 2009).
References Cited:
- Beltrame, L., P. V. Pereira, and L. Berqgvist. 2014. Histological characterization of osteoderms of Tardigrada (Mammalia, Xenarthra) from Pleistocene deposits of Brazil. Journal of Vertebrate Paleontology, SVP Program and Abstracts Book, 2014:89.
- Burmeister, H. 1865. Hauptpanzer bei Mylodon. Archive für Anatomie, Physiologie und wissenschaftliche Medicin 1865:334-336.
- Croft, D. A. 2016. Horned Armadillos and Rafting Monkeys: the Fascinating Fossil Mammals of South America. Indiana University Press, Bloomington, Indiana, 320 pp.
- Gaudin, T. J., and D. A. Croft. 2015. Paleogene Xenarthra and the evolution of South American mammals. Journal of Mammalogy 96:622-634.
- Hoffstetter, R. 1958. Xenarthra; pp. 535-636 in J. Piveteau (ed.), Traité de Paléontologie. Maison et Cie, Paris.
- McAfee, R. J. 2009. Reassessment of the cranial characters of Glossotherium and Paramylodon (Mammalia: Xenarthra: Mylodontidae). Zoological Journal of the Linnean Society 155:885-903.
- Moreno, F. P., and A. Smith Woodward. 1899. On a portion of mammalian skin, named Neomylodon listai, from a cavern near Consuelo Cave, Last Hope Inlet, Patagonia. By Dr. F. P. Moreno, C.M.Z.S. With a description of the specimen by A. Smith Woodward, F.Z.S. Proceedings of the Zoological Society of London 1899:144-156.
- Robertson, J. S. 1976. Latest Pliocene mammals from Haile XVA, Alachua County, Florida. Bulletin of the Florida Museum of Natural History 20:111-186.
- Sinclair, W. J. 1910. Dermal bones of Paramylodon from the asphaltum deposits of Rancho La Brea, near Los Angeles, California. Proceedings of the American Philosophical Society 49:191-195.

You reference Beltrame et al. 2014 in the text but it is missing from your list of references.
LikeLike
Thanks for pointing that out! I will go ahead and fix that.
LikeLike
Boscaini et al 2019 in their paper on the reappraisal of the phylogeny of the Mylodontidae note that osteoderms are only found in the Mylodontini (but not the Lestodontini) and separately in the Scelidodont Valgipes.
So they evolved twice in the Mylodontidae as well as in cingulates.
I wonder if the ancestral condition was to have cartilaginous nodules in the skin of Pilosa for protection and these became ossified to provide extra protection when more large mammalian predators appeared in the Pliocene and Pleistocene. They would certainly mess up the sabre teeth of a Smilodon or Homotherium that made the mistake of attacking them. Their close spacing and unpredictable position would make it highly likely that the tip of a sabre tooth would contact one and get broken.
The living tree sloths could have lost the cartilaginous nodules once they became arboreal due to the lesser rate of mammalian predation in trees; although come to think of it has anyone ever looked for nodules in living sloth skin?
LikeLike
Thanks for you comment.
I think the main drawback of that interpretation is the lack (apparently) of osteoderms in other late Cenozoic mylodontids (and other sloths). If cartilaginous osteoderms were present more broadly among sloths, and if ossified osteoderms were effective in defending against certain predators, it is difficult to explain why they aren’t present in all late Cenozoic mylodontids. The sporadic distribution seems more like a genetic variation that didn’t arise very frequently.
I’m not certain, but I suspect early anatomists took a close look at modern sloth skin and would have noted any cartilaginous nodules. Rob Hill studied the histology of osteoderms in xenarthrans (2006) and non-mammals and probably would have come across any suggestions that something like that might be present in modern sloths. He may have even looked into it himself; I’m not sure.
LikeLike
I checked and you are right, the osteoderms are dermal bones and don’t form from cartilage.
One of the functions of dermal bones is bone respiratory acidosis buffering during prolonged apnea; and it is interesting that cingulates and mylodont sloths are both burrowers and thus could operate in low oxygen conditions in burrows even if not engaging in apnea per se.
LikeLike